Introduction
Stroke is the second leading cause of death and disability-adjusted life-years worldwide [1]. Ischaemic stroke (IS) accounts for 85% of all strokes, while haemorrhagic stroke (HS) accounts for 10% to 15% [2]. Cerebral ischaemia leads to the development of a complex cascade of pathological events centring around neuronal excitotoxicity and adenosine triphosphate depletion [3]. The ischaemic core is surrounded by the ischaemic penumbra, an area of functionally compromised, partially-perfused salvageable tissue destined for infarction without intervention. Elucidation of the mediators of ischaemic cell death within the penumbra represents a promising area for the development of gene-targeted intervention for neuroprotection [4]. Furthermore, the damage that occurs during HS is primarily a result of the rapid accumulation of blood and consequent disruption of brain tissue and increased local pressure. At present, the diagnosis of stroke relies on the clinician’s expertise in patient assessment and rapid access to diagnostic imaging techniques such as computed tomography (CT), CT angiography, CT perfusion or magnetic resonance imaging (MRI). However, clinical assessment of stroke is not always straightforward, since CT images are often normal in early presentation and access to CT perfusion and MRI is still challenging. An accurate, reliable and timely blood biomarker for stroke in the acute setting is currently lacking, although its utility has long been recognised for the purpose of early diagnosis and prognosis, as treatment of acute ischaemic stroke (AIS) is time dependent [5].
MicroRNAs (miRNAs) are short (approximately 20 to 23 base pair) single-stranded non-coding RNAs that play an important role in the post-transcriptional regulation of gene expression through interaction between the miRNA seed region and complementary sites in target messenger RNA (mRNAs), targeting the mRNA for degradation or translational repression [6]. The ability of a single miRNA to modulate the expression of multiple genes in multiple pathways places miRNAs in a powerful position in the regulation of all major cellular functions [7]. The onset of stroke triggers a complex series of physiological, biochemical, and gene expression changes precipitating the onset of neuronal injury, placing miRNAs in an advantageous position in the regulation of multiple neuroprotective and deleterious pathophysiological processes impacting on brain injury maturation. There is a growing body of evidence supporting the potential roles for miRNAs in the attenuation of neurological deficit, modulating key pathways central to the expansion of the ischaemic core. miRNAs dysregulated following ischaemia have been shown to suppress microglial function, playing an important role in the recovery phase by promoting the growth of neuronal tissue, and in the repair of injured brain tissue in the acute phase by stimulating angiogenesis and neurogenesis [8,9]. Conversely, in vitro models have provided significant insight into the potential detrimental effects of miRNAs on signalling contributing to the development of ischaemic injury, such as miR-155 which has been shown to regulate reactive oxygen species production, nitric oxide generation and apoptosis in the endothelial cells [10]. The silencing of miR-155 demonstrates neuroprotective effects in the ischaemic environment by enhancing proliferation, migration, and tube formation in human brain endothelial cells. In the setting of HS, miRNA overexpression has been shown to improve the brain blood barrier integrity, suppress inflammation and inhibit neuronal apoptosis, while other dysregulated miRNAs have the potential to alleviate neurological deficits via suppression of neuroinflammation and reduction of cell death [11-13].
miRNAs represent a novel class of biomolecules with significant attractive properties centred around their ability to target multiple effectors within a signalling network; this is of particular importance given the heterogeneous nature of stroke pathophysiology. Circulating miRNAs are easily accessible and detectable in biofluids, and the identification of unique differential miRNA profiles in blood and urine places miRNAs in a unique position as diagnostic biomarkers for disease progression and measurement of therapeutic response [14-16]. While much effort has been directed towards the identification of protein and metabolic biomarkers, as a result of the low sensitivity of proteins, there is currently no rapid diagnostic assay which would allow better management of acute stroke. The rapid induction of an ischaemic miRNA profile can be detected before the induction of protein markers, and their non-invasive, easy detection by quantitative polymerase chain reaction (qPCR)-based platforms provides rapid and cost-efficient profiling, capable of detecting even low-abundance miRNAs. Another advantage lies in their potential to act as a ‘multifunctional’ assay for diagnosis, informing treatment decisions and evaluating treatment response in the acute phase in contrast to protein biomarkers, where many protein markers would be required, with current analytical methods proving both expensive and time-consuming. While there remains a need for the identification of sensitive blood biomarkers that correlate with the development, diagnosis and prognosis of acute stroke, research centred around miRNAs as biomarkers in stroke is still in its early stages and the findings of such studies generally lack validity. The remarkable stability of miRNAs in biofluids, combined with their ease of access places circulating miRNAs in a promising position as minimally invasive clinical biomarkers in the diagnosis and prognosis of stroke. We here present a comprehensive review of the current clinical status and utility of circulating miRNAs in whole blood, plasma and serum for their ability to diagnose and assist in the prognosis of patients suffering stroke in the acute phase to better inform patient management and outcomes.
Methods
Search strategy
We carried out a systematic review in line with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) reporting guidelines [17]. The protocol was not registered. In order to maximise our search, we combined Medical Subjective Heading (MeSH) terms with keyword terms to formulate the search strings. We conducted a comprehensive search of the databases Medline PubMed, Scopus, Cinahl, Cochrane, Web of Science, and Embase. Our MeSH and keyword search terms were combined as follows: (miRNA OR miRNAs OR microRNA OR microRNAs OR circulating miRNA OR circulating microRNAs OR miR) AND (Diagnostic OR diagnosis OR prognosis OR prognostic OR prediction) AND (Stroke OR acute stroke OR ischaemic stroke OR ischemic stroke OR ischaemia OR ischemia OR haemorrhagic stroke OR hemorrhagic stroke OR cerebrovascular accident OR cerebral infarction OR brain venous infarction). Our search strategy was defined in consultation with a senior medical librarian. All searches were restricted to the English language and no time limitation was applied.
Inclusion and exclusion criteria
We applied the following inclusion and exclusion criteria to our search. Inclusion criteria: (1) case-control studies; (2) image-based diagnosis of acute ischaemic and HS; (3) initial blood samples taken within the first 24 hours from onset of symptoms. Exclusion criteria: (1) animal model studies; (2) non-acute stroke studies; (3) article not available in English; (4) study does not present original data (letters; replies; commentaries; opinion articles); (5) non-blood biomarkers (e.g., cerebrospinal fluid). For papers which were not accessible online, authors were contacted and asked to provide a copy of their study. Papers were excluded from the review where studies were not accessible.
Study selection and data extraction
Following removal of duplicates, paper titles and abstracts were screened to identify relevant papers. Two authors independently applied the inclusion and exclusion criteria to all retrieved citations with arbitration by a third reviewer in the event of lack of consensus. Complete full-text analysis was then carried out on articles that were included following title and abstract screening for final inclusion. Reference lists of identified records were manually checked to identify additional relevant studies. For each study, two reviewers independently extracted and compared data on patient cohorts, sample type and time point from onset, miRNA selection, normalisation and quantification strategies, and 95% confidence interval, area under the curve, sensitivity, and specificity of dysregulated miRNAs.
Included studies quality assessment
The quality of the included studies and risk of bias was assessed by the Newcastle-Ottawa Scale [18]. Studies were scored according to quality of study group selection, comparability, and outcome. A final quality score was calculated for each study ranging from 0 to 9. Studies were classified as high (7 to 9), medium (4 to 6), or low (0 to 3) quality.
Bioinformatic miRNA gene target prediction and analysis
Three miRNA gene target prediction tools (TargetScan v7.2, miRDB [accessed 17th July 2020], and miRWalk v3.0) were used to predict gene targets for miR-16-5p, miR-126-5p, and miR-335-5p, as miRNAs identified as having highest sensitivity and specificity with diagnostic and prognostic potential. Gene targets predicted by at least two of these tools were used for the gene ontology (GO) analyses. All GO enrichment analyses were carried out using EnrichR (version: January 23rd, 2019). An adjusted P-value cut-off of below 0.05 was used to filter results. The code used in the miRNA gene target prediction analyses and GO can be found at: https://github.com/GiantSpaceRobot/miRNA-gene-target-prediction.
Results
Included studies
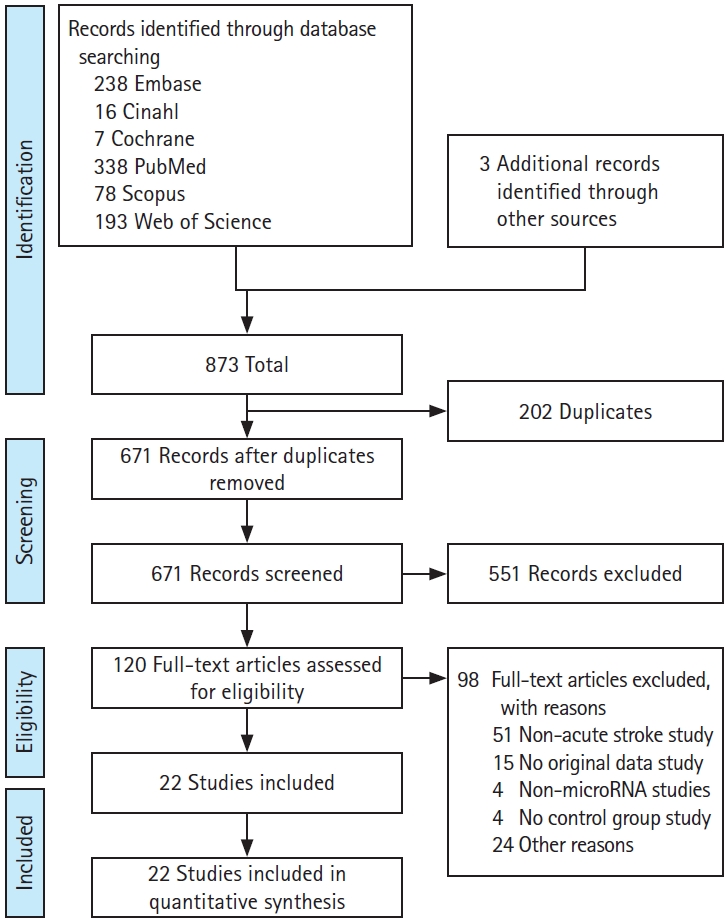
The initial comprehensive search of all the sources yielded 873 papers eligible for screening. After removal of 202 duplicates, 671 unique articles were assessed for inclusion by title and abstract screening, and a further 551 articles were excluded. Complete full-text analysis was carried out on the remaining 120 articles and finally 22 articles were included (Figure 1). Out of 22 case-controlled studies assessing acute stroke, a total of 22 IS and two HS cohorts were assessed, with a total of 2,470 AIS, 69 HS, and 1,651 healthy control (HC) subjects. One study also included transient ischaemic attack (TIA) patients, one recruited non-stroke patients with metabolic complications and one stratified AIS by treatment with recombinant tissue plasminogen activator [19-21]. Five studies recruited less than 50 patients with the remainder recruiting 50 or more [22-26]. Seven studies included only patients with a clinical diagnosis of first IS presentation, excluding recurrent IS patients (Table 1) [22-24,27-30]. Almost all studies excluded patients with a recent history of active infection and active cancer or treatment. Only eight studies classified AIS patients by Trial of Org 10172 in Acute Stroke Treatment (TOAST) criteria which subtypes stroke aetiology into cardioembolism, large artery atherosclerosis (LAA), small vessel disease, and undetermined cause [31]. Included studies did not provide any information about the effect of age and sex on miRNA expression after stroke.
miRNA profiling
Eleven studies assessed miRNAs from plasma [19,21-23,25,27,30,32-35]; seven studies from serum [22,26,36-40]; and four assessed whole blood miRNAs [20,24,28,41]. Six studies assessed plasma-extracted total RNA from blood collected using ethylenediaminetetraacetic acid (EDTA). Six studies utilized miRNA microarray to profile miRNA expression, while only one study employed small RNA sequencing techniques. The remaining studies selected their miRNAs based on evidence from previous work using in vivo or in vitro models or published in the existent literature at the time of the study. A mix of endogenous and synthetic controls were employed for normalisation. Eleven studies used endogenous U6 small nuclear RNA (snRNA), with a further three using RNU44, U75 or a combination of miR-16 and/or miR-451 as endogenous controls. Eight studies used exogenous cel-miR-39, cel-miR-54, or UniSp2/UniSp4/UniSp5 mix (Exiqon) synthetic spike-in controls. Twelve studies employed TaqMan miRNA Assay quantification, while the remaining studies employed SYBR Green-based methods for reverse transcription qPCR (RT-qPCR) analysis (Table 2).
Diagnostic accuracy of identified miRNAs
Diagnostic accuracy of specific miRNAs detected in the acute phase was assessed for sensitivity and specificity (Table 3). We identified miR-335 as demonstrating highest overall sensitivity in AIS (97.6%), followed by miR-16 (94.7%), while miR-16 also showed highest overall specificity (87%) followed by miR-126 and let-7b (84%) (Figure 2A) [25,27,30,32]. Upon assessment of stroke subtype, miR-16 was found to demonstrate the highest sensitivity (100%) and specificity (91.3%) in both large artery atherosclerosis stroke (LAAS) and total anterior circulation infarct (TACI) groups [27]. miRNA-124-3p was reported as having a sensitivity of 68.4% and specificity of 71.2% in HS [32].
Potential clinical utility of dysregulated miRNAs
We define the clinical utility of the reported miRNAs based on their potential to diagnose and predict prognosis in acute stroke. The diagnostic potential was determined by the sensitivity and specificity of the reported miRNAs to correctly identify IS or HS stroke populations, while the prognostic potential was determined by the correlation of the reported miRNAs with clinical outcome such as the modified Rankin Scale (mRS) and National Institutes of Health Stroke Scale (NIHSS) (Table 3). We identified 33 dysregulated miRNAs demonstrating diagnostic potential in the acute phase of the disease (Figure 2B). We identified miR-16, miR-126, and miR-335 as having diagnostic and prognostic utility. miR-16 was reported to be upregulated by two studies, demonstrating both diagnostic and prognostic potential, with the ability to discriminate acute stroke subtype [25,32]. Two studies, Leung et al. [32] and Tian et al. [25], reported upregulation of miR-16 in AIS compared to HCs. Tian et al. [25] reported that miR-16 was significantly upregulated in LAA and TACI subgroups of AIS. Leung et al. [32] reported a correlation of miR-16 with mild stroke, defined as NIHSS <5 but there was no correlation with stroke volume measured radiologically. In comparison to Leung et al. [32], Tian et al. [25] reported that miR-16 expression was significantly higher in the poorer prognosis group (mRS 3-6) than in the good prognosis group (mRS 0-2). Downregulation of miR-126 was reported by two independent groups in three studies to have diagnostic, disease severity management, risk prediction, and monitoring potential [27,34,35]. Jin et al. [35] reported a negative correlation between miR-126 expression and NIHSS score and pro-inflammatory cytokine elevation, placing it as biomarker of disease severity. miR-335 was reported by Zhao et al. [30] as being downregulated and by using multiple logistic regression analysis it is suggestive that miR-335 can act as an independent risk factor for IS. Furthermore, miR-335 correlates negatively with NIHSS. Increased miR-130a was reported as a potential biomarker in early diagnosis of brain oedema and prognosis in HS, correlating with perihematomal oedema severity, NIHSS, and mRS scores [36].
Bioinformatic miRNA gene target prediction and pathway enrichment
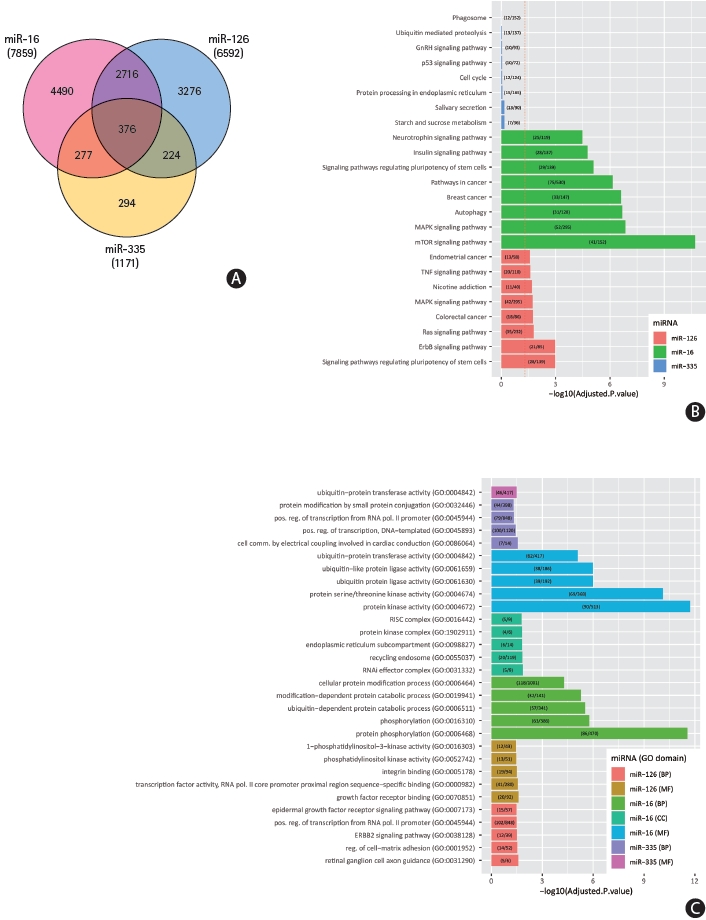
Putative mRNA targets of miR-16-5p, miR-126-5p, and miR-335-5p, identified as having highest sensitivity and specificity with diagnostic and prognostic potential, were identified using TargetScan, miRDB, and miRWalk. A total of 7,859 targets were predicted for miR-16-5p, 6,592 for miR-126-5p, and 1,171 for miR-335-5p. A total of 2,716 were predicted to be shared mRNA targets of both miR-16-5p and miR-126-5p, while 277 and 224 genes were shared targets of both miR-335-5p and miR-16-5p, and miR-335-5p and miR-126-5p respectively. A total of 376 genes were predicted to be targeted by all three miRNAs (Figure 3A).
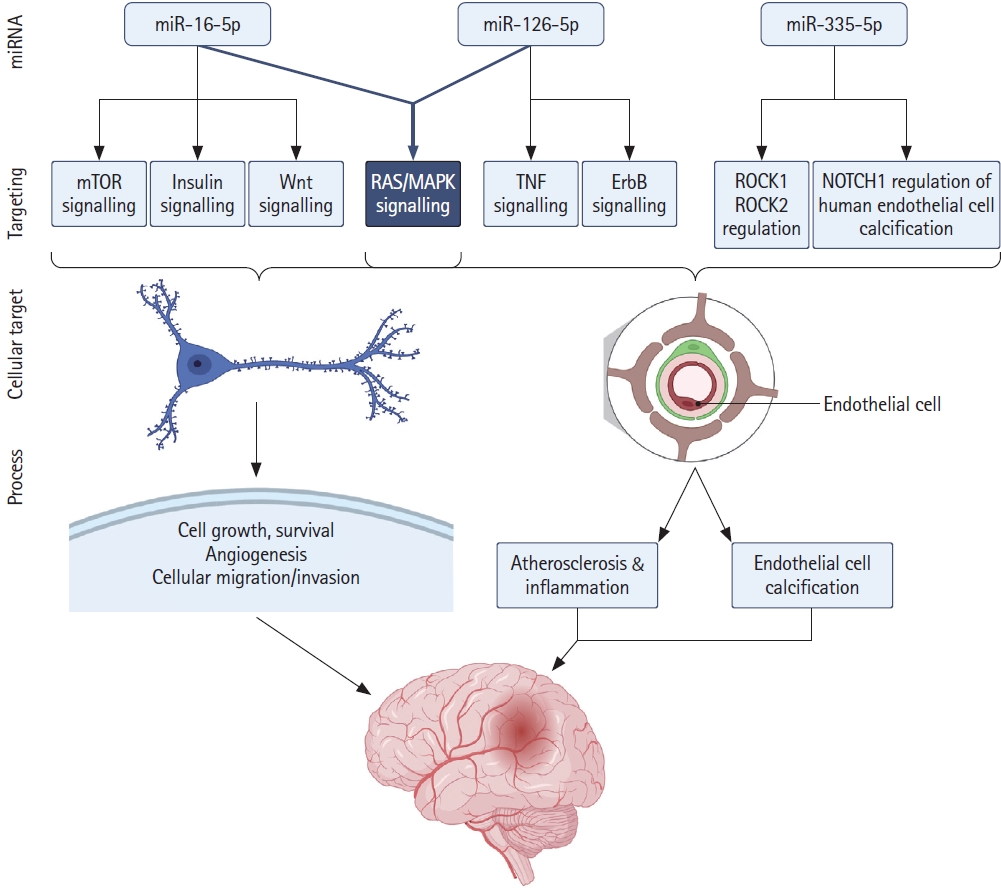
We performed enrichment analysis of GO terms on predicted miRNA-target interactions (Figure 3B and C). The full results are available in Supplementary Table 2 (EnrichR_analysis_Bejleri_2021). Our analysis of miR-16-5p targets revealed significant enrichment (P-adj <0.05) of insulin signalling, Wnt signalling, transforming growth factor β (TGF-β), mammalian target of rapamycin (mTOR), and mitogen-activated protein kinase (MAPK) signalling pathways. Signalling pathways such as these coordinate physiological and pathological processes in response to acute stroke, governing molecular activities at a cellular level. mRNA translation plays a pivotal role in the control of gene expression and regulation of homeostasis in response to disease, tightly controlled by signalling pathways in response to various stimuli, such as oxygen and nutrient deprivation and compromised energy status. Post-translational regulation, degradation or translation inhibition of mRNA highlights the important roles of miRNAs in response to disease pathophysiology. Our enrichment analysis indicates that increased miR-16 in response to ischaemia negatively regulates significantly high numbers of key proteins in the insulin and downstream mTOR and MAPK signalling pathways, impacting on the regulation of critical cellular responses including cellular metabolism, growth, proliferation, and homeostasis (Figure 4). Reported decreased expression of miR-126 following ischaemia carries significant implications for subsequent disinhibition of a significant number of predicted target genes associated with ErbB signalling, tumor necrosis factor (TNF) signalling, RAS signalling, and MAPK signalling pathways. Similarly, downregulation of miR-335 is associated with the positive regulation of predicted target Rho associated protein kinase 1 (ROCK1), involved in vascular inflammation, and NOTCH1 regulation of human endothelial cell calcification, linked to aortic valve calcification and stenosis, placing miR-126 and miR-335 as potential biomarkers of systemic inflammation. Pathways sharing common target genes predicted to be targeted by all three miRNAs are associated with MAPK signalling (13 common targets, P<0.005), ErbB signalling (7 common targets, P<0.005), insulin signalling (7 common targets, P<0.05), and JAK-STAT signalling (8 common targets, P<0.05).
Quality scores
Quality scores for the included studies ranged from 2 to 9 (Supplementary Table 1). Three studies were classified as having low quality as they had a score ≤3 [26,30,36]. Eleven studies were classified as medium quality and eight studies as having high quality.
Discussion
We identified miR-16 and miR-126, reported by more than one study, and miR-335 as having the highest sensitivity as potential biomarkers with diagnostic and prognostic utility in AIS. miR-16 was reported to be upregulated in IS when compared to HS patients and HCs, with particularly high diagnostic capability in LAAS and TACI subgroups. The potential mechanisms of miR-16 in ischaemic reperfusion injury have been extensively studied in middle cerebral artery occlusion (MCAO) mice models and it has been shown that miR-16 promotes ischaemic brain injury by negatively regulating anti-apoptotic Bcl-2 and Bcl-w [42]. Tian et al. [25] demonstrated that miR-16 expression was markedly higher in stroke groups with poorer clinical outcomes (mRS 3-6) compared to those with a more favourable clinical outcome (mRS 0-2).
We report significant overrepresentation of several signal transduction networks by predicted targets of miR-16, notably insulin signalling and associated downstream mTOR and MAPK signalling pathways. Insulin-mediated signalling plays critical roles in the integration of various signals in response to environmental stress and nutrient availability and coordination of cellular response, and downstream activation of mTOR and RAS/MAPK signalling modulates cellular metabolism, growth, proliferation, and homeostasis. Of particular note in the context of our study, mTOR signalling is known to be dysregulated following ischaemia and oxygen-glucose deprivation (OGD), playing roles central to neuronal survival and death, and infarct development. Similarly, activation of MAPK signalling following IS is known to play roles in the regulation of neuronal survival, inflammation, and apoptosis. As a result, these signalling pathways have been the focus of a number of studies attempting to achieve neuroprotection in the treatment of IS, with current evidence indicating that the potential neuroprotective effects of key targets in these pathways do not function in isolation but as part of an integrated signalling response in relation to cellular survival, highlighting the potential importance of the multitargeting regulation of these pathways by miR-16 in the development of cerebral infarction. Interestingly, our analyses also revealed a relationship between miR-16 and Wnt signalling and significant enrichment of miR-16 targets in the TGF-β signalling pathway. Wnt has been shown to be critically involved in neurogenesis and functional recovery following ischaemia while TGF-β plays roles in mediating the angiogenic and immune response in ischaemic brain tissue [43]. miR-16 is highly conserved and ubiquitously expressed across tissues and cell types, with well-characterised roles in the negative regulation of cell proliferation & angiogenesis [44,45]. Increased miR-16 following AIS, negatively regulating complex signal transduction networks identified here, carries significant implications for the regulation of fundamental biological processes critical for coordination of cellular response to ischaemia and associated excitotoxicity. Notably, the critical roles of insulin, mTOR, MAPK, and Wnt signalling have been a focus of significant interest as potential mechanisms affecting cellular survival and injury in the treatment of stroke [46,47].
Three separate studies from two independent groups reported miR-126 as having diagnostic, disease risk prediction, monitoring, and management potential in AIS, demonstrating high sensitivity and specificity alone and in combination [27,34,35]. miR-126 has been shown to be highly abundant and an endothelial-specific miRNA in capillaries and larger vessels, with key roles in vascular integrity and ischaemia-induced angiogenesis [48]. Lower levels of miR-126 have been associated with increased plasma levels of the pro-angiogenic and pro-inflammatory mediator, placenta growth factor, a member of the vascular endothelial growth factor family and primary inflammatory instigator of atherosclerosis development and atherosclerotic plaque instability in coronary artery disease (CAD) and associated disease severity [49]. Decreased endothelial progenitor cell expression of miR-126 has been reported in CAD, heart failure, and diabetic patients [50]. miR-126 has been reported as a potential biomarker of stroke severity as it negatively correlates with the NIHSS, a measure of stroke severity, and its association with cardiovascucardiovascular disease, immune and inflammatory responses. Plasma miR-126 has been shown to negatively correlate with inflammatory cytokine TNF-α, interleukin 1β (IL-1β), and IL-6 levels, while miR-126 over-expression in endothelial cells has been shown to decrease pro-inflammatory TNF-α and high-mobility group box 1 (HMGB1), involved in the pathogenesis of IS and reperfusion injury [35,51]. Our analysis also identified significant overrepresentation of miR-126-5p target genes associated with both TNF and RAS/MAPK signalling, and ErbB signalling, with significant roles, in microglia activation, cytokine production and inflammation, and endothelial cell proliferation and function [52]. Decreased miR-126 and associated positive regulation of target genes in these pathways carries implications for dysregulation of acute inflammatory response and associated stroke pathology, further supporting a possible role for miR-126 as an endothelial cell-specific biomarker of systemic inflammatory status, damage, and severity. This is of particular interest given the strong association of stroke with hypertension and related cardiovascular diseases and associated poor prognosis and mortality.
miR-335 was found to have the highest overall sensitivity (98%) amongst all miRNAs reported, and negative correlation with NIHSS, demonstrating promising diagnostic utility. miR-335 has been reported to be decreased in rat MCAO models of AIS, and synthetic increased expression inhibits apoptotic signalling and reduces ischaemic injury via downregulation of ROCK2 [53]. ROCK1 and ROCK2 are both reported targets of miR-335, involved in fundamental cellular functions including vascular inflammation, cell adhesion, migration, cytoskeleton organization, and stress fibre formation. Our analysis identified ROCK1 as a predicted target of miR-335-5p, and enrichment of miR-335-5p target genes in apoptosis modulation and signalling. We also identified a number of predicted miR-335-5p target genes in NOTCH1 regulation of human endothelial cell calcification. Dysregulation of NOTCH signalling is increasingly linked to aortic valve calcification, a leading cause of heart disease, and dysregulation of miR-335 has also been reported in a mouse model of aortic valve stenosis, supporting a role for miR-335 specificity in patients suffering AIS [54].
Our analysis highlights significant roles for miR-16, miR-126, and miR-335 in the modulation of pathways involved in stroke pathophysiology, immunomodulation and inflammation regulation. It should be emphasized that the functional consequences of miRNA modulation on the identified pathways are dynamic, and both the consequences of the activated signalling pathways and of miRNA regulation of targets are cell- and disease-context dependent. The multi-targeting abilities of miRNAs exert an effect at a systems level in response to environmental and intra-cellular cues, and the impact of target pathway deregulation is dependent on the duration as well as the severity of the insult, such as OGD. Of particular note, we have here identified a significantly high number of gene targets of these ischaemia-associated miRNAs involved in insulin, MAPK, mTOR, and Wnt signalling pathways, carrying significant implications for infarct development and expansion in IS (Figure 4). Elucidation of the mechanisms underlying the pathophysiology of IS represents a promising approach to the development of therapeutic strategies which can halt the progression and expansion of cell death, and promote neuroprotection in acute ischaemia. We identified a small number of common targets of all three miRNAs, involved in the co-ordination of the cellular stress response to ischaemic environment, with impacts on salvageable tissue. Increased sensitivity (100%) and specificity (91.3%) of miR-16 in both LAAS and TACI groups, compared to partial anterior circulation infarct (70.6% and 87% respectively), supports a diagnostic and prognostic role in the severity of compromised tissue. Similarly, decreased miR-126 and miR-335 expression and their association with cardiovascular disorders and regulation of immune and inflammatory responses supports their significant sensitivity and specificity as biomarkers of systemic inflammatory status, damage and severity in IS. In addition to their diagnostic potential in AIS, dysregulation of these miRNAs presents promising utility providing information about underlying aetiology.
Of the identified studies, only one reported significant utility for miRNAs in HS. Wang et al. [55] reported upregulation of miR-130a in haemorrhagic patients and its potential use as a biomarker in the diagnosis of early oedema and prognosis in acute HS. Evidence in both in vitro and in vivo models has demonstrated that delivery of a miR-130a antagonist (antagomir) successfully attenuate brain oedema, reduce both blood-brain barrier permeability and infarct volume, and improve neurologic function [55].
While the utility of circulating miRNAs as biomarkers has been recognised, several factors present problems in their successful translation and development in stroke. We identified wide variation in the number of patients and control groups recruited to each study. However, the diagnostic and prognostic utility of the miRNAs that we have reported were based on robust numbers of recruited patients with a mean cohort 10 number of 74 (range, 38 to 136) [25,27,32-34,36]. Furthermore, 20 of the included studies were performed in Asia, one in Europe and one in USA, indicating that miRNA studies reported may not be representative of all cohorts and population differences in small non-coding RNA expression should be taken into consideration in translational study design [56]. Varied inclusion and exclusion criteria in participant recruitment and sampling time of blood can contribute considerable variability given the rapid evolution of the miRNA profile and the heterogeneity of the developing ischaemic environment. Five studies included only first-time stroke patients or excluded recurrent stroke patients, while four studies excluded patients who had received thrombolytic therapy prior to the blood collection. There is now evidence that tissue plasminogen activator alters miRNA profile expression and subsequently the identification of diagnostic and prognostic biomarkers [21]. All these confounding factors have the potential to impact detection of differential expression of miRNA profiles among the studies and warrant consideration in future study design.
Variability in the methodology used in miRNA studies from blood collection to miRNA profile discovery also warrants consideration. The most common sample type was plasma, followed by serum and a small number of studies employed whole blood. Recent evidence has shown a marked heterogeneity in miRNA expression from serum and plasma samples, impacting significantly on reporting of variances in miRNA profiling [57]. The use of EDTA tubes from six studies adds more potential variability, as anti-coagulants such as heparin and EDTA are potent inhibitors of down-stream PCR reactions, impacting on the quality of down-stream analysis for biomarker identification [58]. These differences in blood collection procedures can affect the coagulation cascade and, along with degree of haemolysis and platelet, red and white blood cell contamination, may influence the biochemical profiles.
Importantly, the most common control for miRNA normalization was endogenous U6, with other studies utilising endogenous miR-16, miR-451, RNU44, and U75. Just eight studies employed exogenous spike-in controls for normalisation. The use of snRNAs, such as U6, may not reflect the biochemical transcription, processing, and expression patterns of other RNA classes and as such may not be suitable for miRNA normalisation. Furthermore, serum levels of U6 have been reported to demonstrate a high variability in patients with critical illness, and to correlate with serum levels of established inflammatory biomarkers [59]. Use of circulating miR-16 and miR-451 as a normalisation strategy, known to be significantly altered in plasma and serum in response to haemolysis, for blood samples from patients with acute intracerebral haemorrhage by one study in this review highlights the problematic area of lack of consensus on a normalisation strategy, particularly in the case of a haemolytic-susceptible circulating diagnostic or prognostic biomarker such as miR-16. However, due to the sensitivity and specificity of miRNA profiles in response to disease states, especially given the heterogeneous nature and aetiology of IS, the identification of a single reference with universal applicability will be difficult. To this end, the use of exogenous synthetic spiked-in miRNA during RNA isolation for normalisation represents a robust approach.
The considerable variation in the pre-analytical and analytical approaches highlighted here represent a significant barrier in the identification of clinically useful miRNAs in translational stroke research, and are most likely responsible for the heterogeneous results yielded from our analysis and the number of studies that have reported the top three miRNAs which we have identified as having diagnostic and prognostic potential. The small number of studies which report these miRNAs raises concerns regarding the reproducibility of such studies. However, our bioinformatic analysis has demonstrated the pivotal role that these miRNAs play in stroke pathophysiology. The variability across profiling platforms, and in particular the lack of consensus on normalisation strategies confers a significant impact on the ability to detect significant biological differences across samples is such a heterogeneous disease setting, and therefore their utility in a clinical and research setting. Nonetheless, we present a comprehensive and encouraging review of current evidence on the current position of miRNAs as biomarkers of acute stroke and our analysis indicate the need for further research in order to confirm our findings and provide more evidence of the potential clinical utility of miRNAs in the field of translational stroke research.
Conclusions
We identified candidate miRNAs miR-16, miR-126, and miR-335 as promising biomarkers for stroke with diagnostic and prognostic potential. Considerable variation in pre-analytical and analytical parameters which potentially contribute to the lack of overall consensus in miRNA expression profiles identified in this review highlights the need for internationally accepted and unified methodologies which would standardise the process of blood sampling, handling and preparation for miRNA screening, profiling and validation. It is important to acknowledge the potential contributions of underlying pathologies to miRNA dysregulation in the setting of AIS, as we have highlighted in our bioinformatic analyses. Inflammatory response has been well established as a biomarker of stroke, and miRNA changes may reflect the underlying aetiology and systemic inflammatory responses as factors related and consequential to AIS pathogenesis. While considerable advancements have been made in the development of miRNAs as therapeutic targets in non-stroke pathologies, the heterogeneity of stroke pathogenesis and associated biochemical cascades requires more robust investigation for the meaningful translation of miRNAs as therapeutic modulators in AIS. While the capacity of miRNAs to target multiple genes in multiple pathways places them in a unique position for development in AIS, the complexity of stroke pathophysiology and associated, equally complex, comorbidities requires greater consideration in the development of candidate miRNAs as biomarkers and potential therapeutic targets. Consideration for toxicity and off-target downstream effects resulting from the delivery of miRNA mimics and antagomirs highlights the need for robust pre-clinical development for the refinement and successful translation of miRNAs as multi-targeting biomarkers and promising therapeutic targets in AIS.